2. ユダヤ人周辺のDNA
(1) ユダヤ人固有の DNA:
ユダヤ人とは、すでに多くの人種が混合してしまい、現在では ユダヤ教を信奉する宗教的ユダヤ人が”ユダヤ人”と定義されている。それにもかかわらず、アシュケナジー、スファラディーを問わず、コーヘン(男系で継承する 祭司職の家系、 イスラエル12部族の「レビ族」よりも狭いグループ)にだけある特徴的なDNA断片群が見出され、ユダヤ人種を識別するのに用いられている。
* アシュケナジム: 東欧系ユダヤ人・アシュケナズ=ドイツの意、 現在 米国のユダヤ人の大半)
セファルディム: 1492年に追放されるまでスペインに住んでいたユダヤ人で、その後北アフリカなどに拡散、元 駐日大使のエリ・コーヘン氏など
ミズラヒム: 長らくアラビアに居住、 ミズラ=東
その他、 エチオピアのダン族、 ミャンマーのマナセ族、 ジンバブエのレンバ族 など。
1997年、テクニオン-イスラエル工科大の腎臓の専門家であるカール・スコレスキー(Karl Skorecki) 博士と、アリゾナ大学の遺伝学の専門家マイケルF. ハマー博士によって、その特徴あるDNA配列がY染色体上にあることが発見された。
Y染色体のイントロン(ジャンク(がらくた)遺伝子 = 生体機能を発現しない部分、全DNAの95%以上)における、突然変異が起こった場合その形質が継承されやすい3つの部位(サイト)と、突然変異が頻繁に起こる6つの部位(サイト)の、 計9つのサイトを一つの組として調査し、コーヘン階層では非常に多く変異が起こっているが、普通のユダヤ人の間ではそれほど一般的でなく、ユダヤ人でない集団の間では稀であるという結果が得られた。その 9つのサイトがすべて変異している割合は、
アシュケナージ系コーヘンで45%、 スファラディー系コーヘンで56%、 一般のユダヤ人では全体の3~5%
したがって、アシュケナージ系コーヘンも、スファラディー系コーヘンも、ほとんど同じように約半分はこのコーヘン特有の男系遺伝のDNA群を保っていることになる。(それぞれ1000人近くからサンプル) アシュケナジーを にせユダヤ扱いする人もいるが、実際はそうではなく、ロックフェラー、ロスチャイルドのほうが DNA的に偽者であることがわかる。(→ 下のJ2a系統参照)
この調査結果により、アフリカ・ジンバブエの レンバ族がユダヤ人の末裔であることが証明された。 彼らは、外見上完全な黒人であり、割礼と安息日を守り、豚肉とカバの肉を食べないなどの風習を持ち、イスラエルからセンナという所を経て南アフリカに移住してきた という伝承が代々伝えられている。また、彼らの祭司職のグループは”ブバ族”と呼ばれ、彼らによるとブバ族はイスラエルの12部族の一つであるとされている。彼らのY染色体DNAの変異箇所の調査により、レンバ族全体の男の 9%、”ブバ族”では 53%が、コーヘンに特徴的なDNA変異を持っていることが分かった。これは、イスラエルやアメリカのユダヤ人と全く変わらない割合である。(by.ニューヨークタイムズより、 文献1999年、 ニューヨークタイムズ(2003 9 27・再))
注) アシュケナジー系は ハザール国(コーカサス)とは全く無関係。(↓(2)の2)) またアシュケナジーはれっきとしたユダヤ人であり(↑)、東欧の血が濃い。(↓(2)の4)) これに対して、”にせユダヤ”とは、ワスプ(ドイツ系白人)のロックフェラーや クレタ島出身のロスチャイルドを言う。
ただし、北イスラエル10部族のDNAについては、祭司職を含む「レビ族」の大部分が南ユダと北イスラエルの分裂の時、ヤロブアムから職を解かれ、南ユダへ移動したので(Ⅱ歴11:13-15)、今のところ、このDNA変異箇所によっては、イスラエル10部族の手がかりを知ることはできず、新たに特徴あるDNA変異を見つけなければならない。その中にまだユダ族などが混じっていれば、ユダと共にいるコーヘンのこのDNA断片群が発見されるかもしれない。
* コーヘンのY染色体DNAのハプロタイプは、 J1a1b1b1a2a1a1a4(J-L147.1*, subclade-YSC234) ・・・・ (J系統のハプロタイプ) で、紛れも無くJ系統。ただし、J1?(マホメットなどと同様に、むしろアラビア人に近い??) 他の一般的なユダヤ人は、J2a系統。(↓)
** 英語版ウィキの『アロンのDNA』では、 コハニムの約半分が J1c3(J-P58)、 15%が J2a(J-M410) となっている。(Y-chromosomal Aaron) J系統 というだけなら、アシュナジー・コハニムの 88%(67人、n=76)、スファラディー・コハニムの 76%(52人、n=68)で、どちらも8割は J系統。 残りは E3b、R1b、R1a、K2など数%。
・・・・ 確かにモーセ五書におけるアロンの出自には曖昧さが残る。「レウィイ(レヴィ)族のあなたの兄弟アハロン(アロン)」(出エジプト4:14)で登場し、エジプトにおける在留異国人のイシュマエルの子孫の可能性がある。(兄弟 アフ(EX) = 創世記25:18「(イシュマエルの子孫の)すべての兄弟たちは互いに敵対して住んだ。」: 肉親の兄弟のほかに、民族的な兄弟の意味もある) モーセが口下手ではなくアロンを必要としなければ、J2aだけだったかもしれない。 モーセは、アムラムとヨケベデ(どちらもレヴィ人)の子供。(出2:1、民26:59) アロンの子の祭司はエルアザル。大体ここから、すべての祭司職への父系遺伝が繰り返されていった。そして、意外にも、長い歴史の中で比較的よくその特徴あるDNAが保たれてきたことが立証された。
(2) ユダヤ人が属する Y染色体DNAハプログループ: Y染色体DNAハプロタイプ系統樹(2003) 、 List of haplogroups of historic people(有名人のハプロタイプ)
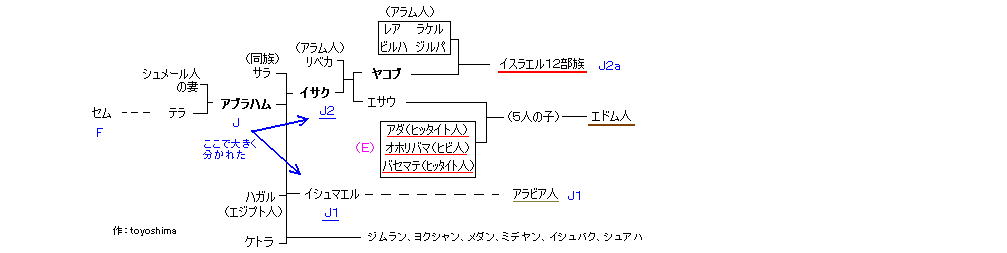
F(セム) > G、H、I、 J > J系統=アブラハムの子孫 > J2a が オリジナルのユダヤ系、 G2c(アナトリア) は捕囚の時混血? or ディアスポラ時代?
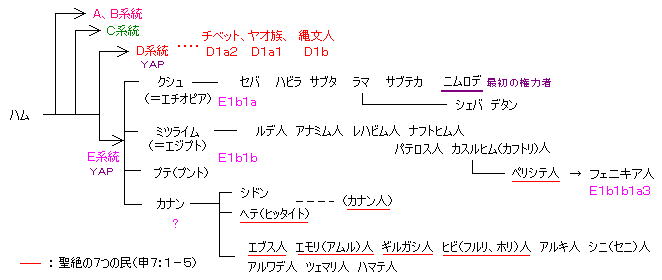
(ハム) > A、B、C、 D、E > E系統= クシュ、ミツライム、プテ、カナン > E1b1b1a3 は 主にフェニキアとの混血
K(ヤペテ) > L、M、N、O、 P > Q、R > R1a1 は ディアスポラの時に東欧系が混血 (アシュケナジムのみ)
1) J2a系統: Y染色体DNAハプロタイプの J系統は、セム系の J1系統と J2系統に分かれる。(ヨーロッパのY遺伝子分布)
・ J1(M62、中東の南部系) ・・・ アラビア半島全体 30%、 特に イエメン 70%
ex) モハメッド(イスラム教創始者ムハンマド、570-632; ヨルダン王族2名より分析) J1a2b2*、 ペルシャ・カージャル朝の王(1772-1834) J1
・・・・・ これはどう見ても イシュマエルの子孫である。父親はアブラハムで、母親はエジプト人。
・ J2(中東の北部系) ・・・ J2a(M41): クレタ島、インドのバラモン階層(R + J2a)
J2b(M63): ギリシャ、アナトリア(トルコ中部)、バビロニア、エルトリア(ローマ初期の支配層)
この J2a 系統に ユダヤ人が集まっている。 J系統は アブラハムの子孫と考えられ、オリジナルのヘブル民族(セム → エベル)の流れがここにある。
∴ J1: イシュマエルの子孫(ハガルはエジプト人)
J2: イサクの子孫(=イスラエル民族、サラは同族) + ケトラの子孫(ミデヤン、 ヨクシャン → シェバ、デダン など、 創25:1-4)
ex) あるロシア系ユダヤ人 J2a1b(J-M67)、 あるルーマニアのユダヤ人 J2(J-M172)、
ロスチャイルド家 J2a1b3(J-L210) ・・・ もちろん、J2aであるからといって本当のユダヤ人であるとは限らない。(ロスチャイルドはJ2a(M41)の多いクレタ島出身であり、実は にせユダヤである。ユダヤ教は全く信奉していない。 「クレテ人(クレタ島の人)は昔からのうそつき、(性的な意味で)悪いけだもの、なまけものの食いしんぼう」(テトス1:12) ・・・ BC600頃の クレタ島出身の哲学者エピメニデスの格言を 流用。当時、キリスト教宣教を妨害する 評判の悪い自称ユダヤ人たちがいて、間違った教えで人々を翻弄していた。
(ロックフェラーはドイツ系白人のワスプで、出自が明らかに異なる。デビッド・ロックフェラーのユダヤ教は形だけ。)
ヨーロッパのユダヤ人家系は J2が多い。現在のイスラエル国はJ2が 20-30%。(レバノン 30-40%、ヨーロッパ 5-10%、その他 アナトリア、イタリア中部、南イタリア、アラビアなどに広く分布) ところが コハニム(レビ族のうちの祭司職、アロンの男系子孫)に アラビア系の J1系統が集まっているのは驚きである。主が しるしのために、本来のユダヤ人でないアロンを任命したとしか考えられない。(↑の(1))
また、ヨセフの時、外国人支配時代のエジプト(ヒクソス時代)なので、エフライムとマナセには E系統(YAP)は入っていないで J系統・セム系が保たれていると思われる。
晩年のアブラハムのケトラの子(東に行ったが)は 父系遺伝から J系統であり、日本人にはJ系統がいないため、日本人がケトラの子孫であるという説は間違い。
2) G2c系統: これもセム系で、アシュケナジ系ユダヤ人に G2cのみが見出される。コーカサスのG2aは入っていない。
・ G2a ・・・ コーカサス(黒海、カスピ海の北)、 イラン、アフガニスタン、カシミール
ex) ヨシフ・スターリン G2a1a、 アイスマン(コルシカ島・特殊) G2a2a1b
・ G2c ・・・ アナトリア(トルコ中部)、 アシュケナジム・ユダヤ
・・・・・ したがって、アシュケナジムが ハザール王国(コーカサス)の改宗者の子孫(7-10c)という説は、DNAから見て間違いで、現在では俗説とされている。
3) E1b1b1a3(E-V22)系統: E系統は、創世記10章の ハムの子孫(クシュ、ミツライム、プテ、カナン)の代表であり、D系統と共に、Y染色体DNAに YAP変異を持ち、アフリカ、地中海沿岸に、ごく普通に非常に幅広く分布している。(A系統、B系統を半ば駆逐するような形で) 日ユ同祖論に近い議論でたびたび言及される YAP共通の点は、E系統と D系統が ごく初期の段階で分かれ、その後は互いに交流の形跡が無いことから、YAPのしるしはそれほど重要な意味を持たないと考えられる。(初期の 階段状ピラミッドまでは共通と思われる。 バベルの塔 → エジプトの階段状ピラミッド → エジプトの大ピラミッド → 小規模のピラミッド状の墓; → チベット・中国のピラミッド、 → 日本の海底ピラミッド)
・ E1a(E-M33)系: 西アフリカ (→ るいネット)
・ E1b1a(E-M2) ソマリア、エチオピア、ギニア、サハラ以南 ・ E1b1a7(E-M191) モザンビーク、ジンバブエ、南アフリカ
ex) ラムセス3世 E1b1a(E-V38) (・・ 2012 12)、 バラク・オバマ E1b1a(E-V38)
・ E1b1b(E-M215、M35 (旧)E3b)系: 東アフリカ、北アフリカ、南ヨーロッパ(ドイツとオーストリアに9%弱もいる)
ex) アドルフ・ヒトラー E1b1b1(E-M35) ・・・ E1b1b1a3 と異なり、フェニキア・ユダヤ系ではない
・ E1b1b1a(E-M78、 (旧)E3b1a)、 ・ E1b1b1a1(E-V12)、 ・ E1b1b1a2(E-V13) 古代ギリシャ人、 ・ E1b1b1a4 モロッコ人
・ E1b1b1a3(E-V22) フェニキア人、古代ユダヤ人(レヴァント(=シリア、パレスチナ(ペリシテ人の地)))、アラビア半島
・・・・・・ E系統のユダヤ人の多くは、初期の歴史の中で本来のJ系統ユダヤ人グループに合流したものと考えられる。 ソロモン以降、フェニキアとのかかわりの時代に 積極的な交流があったことを示す。(アハブの妻イゼベル)、 さらに古くは ヒッタイト(ヘテ人)、モアブ人などが混じっているはずである。
● ハム系 → ミツライム(エジプト) → カスルヒム → ペリシテ人(E系統) + J2系統(シリア人) ≒ フェニキア人(言語はセム語系の フェニキア語、 人種は混合) → BC9cにはアッシリアに服属、 カルタゴへ移る → ポエニ戦争以降 共和政ローマに併合
ハム系 → シドン人
ハム系 → カナン → ヘテ人(ヒッタイト) ・・・ エジプトに、”鉤鼻”のヒッタイト兵士の彫刻、ヘテ人とはアブラハムの頃に交流、
エサウの妻(ヘテ人2人と ヒビ人1人) → 5人の子 → エドム人 ・・・ このエドム人がフェニキアの航海技術を持ち、ユダヤ人と共に 南洋ルートで沖縄・山陰に来たと考えられる(→ (1) エドム人とユダ族の移動 ) 琉球人から見たフェニキア人は 背の高い人々だったとされる(下に 沖縄のフェニキア遺跡)
モアブは、アモンと共に、アブラハムの甥のロトと彼の娘たちとの近親相姦の子孫
・ E1b1b1b2(E-Z830) アルバート・アインシュタイン ・・・ 上記(E1b1b1a3)と異なるので、本当のユダヤ人ではない
・ E1b1b1b2a1*(E-M34) ナポレオン・ボナパルト ・・・ 家系が中部イタリア出身
・ E2(E-M75) サハラ以南
・ セファルディム(北アフリカ)や、特に エチオピアのユダヤ人(ベト・イスラエル(ダン族)、A3b2(M13) 41%、E3*(M40) 18%、E3b(P2) 32%、 J2(M172) 5%・・・ E系統50%、黒人系で90%以上、ユダヤ系は5%もいない)への E系統の混入率は高い。(ディアスポラ時代に混入)
4) R1a1系統: R1a1: 東欧系、 R1b: 西ヨーロッパ人の40~80%
2005年の調査により、 アシュケナジー系男系 Yの 11.5%が R1a1系統であり、母親がユダヤ人で 一部の東欧系の父親の子供がユダヤ教になったことを裏付けている。 ( R1a: 東欧人で 54-60%、2001年) 母系(ミトコンドリア↓)の80%以上との違い
* また、アシュケナジムのミトコンドリアDNAの解析では、はるかに多く、母系の81%がヨーロッパ系(mt-DNAの H 20.5%、K 31.8%、N1b 9.2%、J6.3% など)、中近東(ペルシャ、アナトリア) 8.3%、アジア1.1%であり、80%以上が(コーカサスではなく)ヨーロッパ人種であることを裏付けている。(→ Nature Asia、 Figure10 (2013 10))
これは、エルサレム陥落後まもなく ヨーロッパ人の妻を多く娶ったためといわれる。歴史では、8-9世紀に北フランスにユダヤ人集団の記録があり、14末-15世紀にかけて西ヨーロッパを追放され(イギリス1290-、フランス1394-、ドイツ15c-)、東欧へ移動し、特に ポーランドの保護に入った。この時、特に 現地人の妻との間に子孫が多くできたことを表している。(同族結婚では奇形などが多くうまく育たなかったと思われる。)
** アシュケナジー系の 人口ボトルネックの存在:
25~32世代前、すなわち 1200-1400年頃に アシュケナジムが わずか250-420人程度にまで減少した形跡が発見された。これは、ヨーロッパ人と比べて、劣性遺伝疾患が100倍に増幅されていることなどから推測されたものである。 この時期には、十字軍による組織的虐殺、あるいは ペストの罹患 および その迫害 という、きわめて厳しい環境下に置かれた歴史を経てきた事を表している。(申命記28章の のろい、「星の数ほど多くても、残された者はわずか。」(28:62) ただし、今は回復の時になってきている。)(「東欧系ユダヤ人は250~420人が創始者」ゲノム解析で判明、2014、記事元・WIRED.jp.)