5. 遺伝メカニズムの考察
(1) 個人と集団の基本的な遺伝特性:
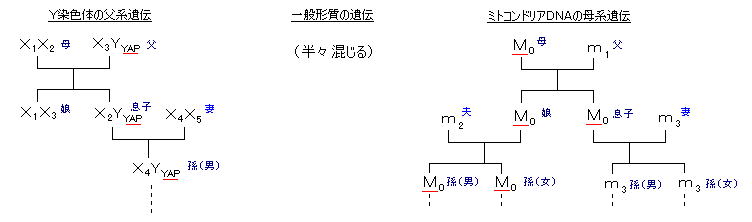
細胞の呼吸をつかさどるミトコンドリアに含まれるDNAは、塩基総数が約16500、測定されるのはその一部(Dループ: 1100個の遺伝に関わらない塩基列部分(=”がらくたDNA”)、変化が激しいので個人差がよく現れ系統樹が作成できる)のさらに一部(482個、190個などの塩基列)で、それぞれミトコンドリアDNA全体の1~3%程度。
父親の精子によって100個程度のミトコンドリアが受精卵に持ち込まれるが、それにはユビキチンというたんぱく質がついていて、その目印のついたミトコンドリアを選択的に破壊するメカニズムが知られている。(時々破壊されないで残る場合がある) したがって、ミトコンドリアのDNAは母親からのものであり、さらにそれは祖母からのものであり、曾祖母から・・・のものであると辿っていく事ができる。(母系遺伝) したがって、あらゆる人の”母方”は一人の女性に行き着く。(ウィルソンのミトコンドリア・イブ) その遺伝の途中で、突然変異が起こっている(DNAの何個かが入れ替わっている)ならば、入れ替わった地点が「分岐点」であり、人種の分類を決める系統樹を形作ることができる。
ミトコンドリアDNAの調査結果によると、日本人の10数%については、母方がアフリカ・中東系に近い人からのものであることが”しるし”として残っているのである。
また、一般の形質は、父と母からおおむね半々を受け継ぎ、ある人の遺伝子の各部分はそれぞれ何代か前のある祖先からのある部分が発現していることになる。したがって、たとえば一人のアフリカ人の父や母が祖先の中にいたとしても、アジア人の血が次第に濃くなるならば、形質は全くの”アジア人”になってしまう。形質の一つである血液蛋白の分析では、アジア6集団(韓国人、モンゴル人、本土日本人、チベット人、アイヌ人、沖縄人)はヨーロッパ、アフリカなどに比べ非常に接近していた。他の形質についても差は緩やかにしか現れない。(・二重まぶた:アイヌ人96.8%、朝鮮南部42.1%、・アフリカ南部のレンバ族のY染色体のDNAには、レビ族(祭司階級(コハニム)、アロンの子孫)に特有の塩基列があるが、外見上は完全な黒人である。)
したがって、一般的な形質については、集団間の相対的な遺伝距離しか知ることができない。
さらに、Y染色体の遺伝子は、もっと単純なメカニズムによって父系遺伝する。約1/2の確率でXYに結合した受精卵は、Y染色体上にあるSry(スライ)遺伝子が身体が男性になるように指示する。核DNAの塩基数は約60億個で46本の染色体に収められている。(ミトコンドリアの塩基数の約40万倍) このうち、Y染色体のDNAは6000万塩基であり、しるしとなるYAPはそのうちのわずか300塩基の部分である。常、Xに比べ、Y染色体DNAは機能を持つ遺伝子が極めて少ない。 当然、Y遺伝子の調査対象は男性のみであり、男性にいわゆる”縄文系”、”弥生系”の区別があり、女性にはこの区別がない。(cf. ミトコンドリアは男性も女性も持ち、しかも母系遺伝する。)
ただし、Y染色体遺伝子は、しばしばDNAが欠落し正確に伝わらないことがある。
(2) Y染色体DNAによるマナセとエフライムの考察:
Y遺伝子DNAによる世界の分布と、アミシャブによるマナセのいる所の調査結果から、このエフライムとマナセの2部族だけがD系統の父系遺伝を持つらしいことが分かった。
このことには、次の5つの可能性が考えられる。
① エフライムとマナセの元の集団が純粋なヨセフのY遺伝子を持っているとして、南北朝分裂の後、イスラエル近辺にいた偶像崇拝のハム系の民(カナン人、ペリシテ(=パレスチナ)人、フェニキア人など)の父方集団の血がかなりこの集団に混合し、そして皮肉なことに、そのハム系のDNAが逆に、エフライム族、マナセ族のしるしとなっているという見方である。(ホセ7:8-16などの記述参照)
E系統は、アフリカにきわめて多い(モロッコ80%、南アフリカ50%、中央アフリカ・ピグミー60%、カメルーン70~80%)ので、純然たるハム系である。また、古代ヘブル人のDNAは、レビ人以外は明確になっていない。このレビ人にもかなり混じっている。(エチオピアのユダヤ人E:32%、D:0%)
しかし、アイヌ人のようにD2の比率が88%と、混じったにしてはあまりにも高率であり、また、日本人にE系統が見られないこと、地中海沿岸や他のアフリカ地域にD系統が全く見られないこと、同様に姦淫したであろう他のイスラエル8部族や南の2部族(ユダ、ベニヤミン)からD系統が見出されないことなどから、この考えは即受け入れることはできない。姦淫は、イスラエルの国内や部族内のみで行なわれたと思われる。ホセア書の記述は、Y染色体の遺伝子ではなく、一般形質の遺伝子について、この時、また、後の日本での時代にも混じったことを意味する。
* レバノン(セム言語系) E26%、F16%、J29%、R16%(31人)、
シリア(セム言語系) E20%、F30%、J15%、R25%(20人)、
トルコ(アルタイ言語系) E16%、J40%、R14%、他G、Iなど(30人)、
グルジア(カフカス言語系) E0%、G30%、J33%、R28%(63人)、 などとなっていて、D、DE*系統は無い。
② それ以降のアッシリア捕囚のとき、同じアフガニスタン北部にD系統の別の民族がいっしょに強制移住させられ、その後、エフライムとマナセと移動を共にしたという考え方。これも、現在のパレスチナやアフガニスタン近辺などの西アジアにD系統の民族が全く存在しないことから考えにくい。
③ また、イスラエルの民が400年間のエジプト滞在中、及び、出エジプトの時に、エジプト人の血が混じったという考え方。これも、エジプト人がイスラエル人とともに食事をすることさえ忌み嫌っていたこと(創43:32)、エジプトでイスラエルが別の居住区に住み、奴隷として使われていたことなどから、それほど多くないと考えられる。出エジプトの民の中に、在留異国人(エドム人(エサウの子孫・毛深い)、エジプト人)やエジプト人を父、イスラエル人を母とする者が一部いたことは確かである。(申23:7、レビ24:10)
④ 日本人のD系統が、そもそもエフライムなのではなく、シュメール人(バビロニア、シヌアルの地 ; ハム系)が出身母体で、その地の民は滅びてしまった、という考え方。シュメール人も、歴史の中に忽然と現れ、また突然消えた民である。しかし、古代日本のイスラエル性だけでなく、同じD2系統のチベット近辺の民が、アミシャブが認めているマナセ族の末裔であることから、D2が文化的なイスラエル性を持つ人種であり、歴史の終わりに5000もの神々を持つようになったシュメール人が日本人であるとは考えにくい。(宗教的には、アイヌ人はその後北方アジアの影響を受けてアニミズムに変わり、本土人は渡来人による影響を受けて仏教になったと考えられる。) また、縄文人の中に、シュメール人特有の”楔形文字”や”青銅器”などの文化は全く伝わっていなく、ゆるやかな移動の途中と思われる道筋(南アジア・ルート、ユーラシア大陸)にもD系統やそれらの文化は見出されない。
(* 戦前の”シュメール-日本人説は、戦時下の日本人の士気を鼓舞し、日本人の優越感を植え付けるために吹聴されたものに過ぎない。)
また、アブラハムがカランを出る時、「カランで加えられた人々を伴っていた。」(創12:5) もし、彼らシュメール人がD系統の特徴を持つならば、その特徴はイスラエル12部族のすべてに行き渡っているはずであるが、他の10支族やアラビヤ人(アフロ・アジア語系、イシュマエルの子孫=母ハガルはエジプト人(創25:12))などにはこれがない。(E系統は存在する)
残る可能性としては、次のようなものである。
⑤ やはり、マナセ、エフライムが特別なD2系統であるという説は捨てがたい。状況としては、D、E系統は昔、共通祖先から分岐したはずであるにもかかわらず、現在はアジアの東西に偏極し、地理的にD、Eが同時に存在して重なる地域がない。(ウズベキスタンにはわずかに両方ある) しかし、D2の多い日本本土・琉球とチベット近辺にのみ、それぞれエフライム、マナセとしての古代ユダヤの伝統が受け継がれている。さらに、日本、チベット、東アジアには、ユダヤ性を伝えたと考えられるセム系らしい系統(F、G、H、I、J)が全く存在しない。(ただし秦氏の多い近畿・関西などは、現在のところ調査データが無い) したがって、D2を持ち、なおかつ、ユダヤ文化の独自性を持った大きなグループが、短期間で東の方へ移動して、朝鮮半島から日本に入って住みついた、としか考えようがない。
系統の分岐はDNA配列の突然変異によって出来、しかも、DE*祖型からDとEへの分岐はかなり昔の段階にある。(DE*祖型はナイジェリアに存在するらしいことが5例報告されている)
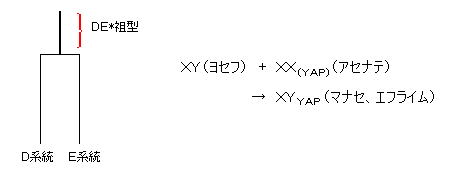
すると、12部族のうちのエフライムとマナセだけがYAPなどの挿入部をもつD系統であることから、ミトコンドリアDNAの結果の考察と同様に、集団ではなく昔の個人、すなわち、エジプト人であるヨセフの妻のDNAが突然変異したことが考えられる。では、どのようにして突然変異したのだろうか?
精子(XY)と卵子(XX)が受精する時に起こりやすい突然変異を考える。
一つのDNA塩基が別の塩基に変化する”置換型”突然変異よりも、すでに存在するDNA配列の”挿入”や”不等交叉”の突然変異の方がはるかに起こりやすい。たとえば、色覚異常について、X染色体上の赤オプシンと緑オプシンの遺伝子は近接して位置しており、DNAの配列も似ている。(”遺伝子重複”によって作られたと思われる) これらの遺伝子は時々混じり合い(不等交叉)、遺伝子の配列がモザイク状になって、色覚異常を引き起こす。
もし、YAP+という、DE*、D、E系統を特徴付ける塩基配列がヨセフの妻(ハム系)のX染色体のどこかに隠れて存在していたならば、それが受精の時に、ヨセフ(セム系)のY染色体のDNAの中に”挿入型”突然変異として入り、息子(マナセ、エフライム)のY染色体DNAはD系統に変化してしまった(ここから、D系統がスタートした)と考えられる。
XY(ヨセフ) + XX(YAP)(DE*:アセナテ) → XYYAP(D:マナセ、エフライム)
現在、エフライムを自称する民族は多いので、神様は、この特別な変異のしるしを エフライムとマナセだけに与えたのではないだろうか?
このような特別な遺伝をした場合、アセナテからのハム系の遺伝子が入ったのは1回きりだから、一般形質としてはセム系の形質がある程度は残っていると考えられる。(・彫りの深い顔、・毛深い、・長い手足、・低い身長 ・・・ cf. ハム系のカナン人は背が高かった(民13:32-33))