丂丂丂丂丂丂丂丂丂丂丂俁丏丂僲傾偺峖悈偺梫慺偵傛傞擭戙廋惓 偲 屆戙擔杮恖偺俢俶俙
丂丂乮侾乯丂僲傾偺峖悈偺梫慺偵傛傞擭戙廋惓偲撽暥帪戙丗
丂丂扽慺侾係朄偵傛傞擭戙應掕偼丄偒傢傔偰旝検偺曻幩惈扽慺侾係乮尰嵼偺擹搙丗俠14/俠12亖侾丏俀亊侾侽-12乯偺應掕側偺偱丄應掕偵崅搙側媄弍偑梫傞忋偵丄應掕岆嵎偑偐側傝戝偒偄丅乮懠偺曻幩惈暔幙偱偼丄應掕抣偲偟偰慡偔巊梡偱偒側偄傎偳岆嵎偑戝偒偄乯丂偦偟偰壗傛傝傕丄扽慺侾係偺検偑戝愄偐傜曄傢傜偢堦掕偱偁傞偙偲傪慜採偲偟偰偄傞丅
丂丂偟偐偟丄懡偔偺徹嫆偵傛偭偰丄峀斖埻側戝峖悈乮僲傾偺峖悈乯偵傛傞抧媴婯柾偺寖曄婜偑懚嵼偟偨偙偲偑柧傜偐偵側偭偰偄傞丅丂偙偺戝曄壔偵傛偭偰丄戝婥偵偁偭偨乽戝偄側傞悈偺憌乿偑徚柵偟偰尰嵼偺傛偆側戝婥寳峔憿偵側傝丄朞榓嬋慄揑偵扽慺侾係偺検偑憹壛偟尰嵼偺斾棪偵側偭偨偲峫偊傜傟傞丅乮扽慺侾係偺検偼尰嵼傕憹壛拞偱丄崱偐傜栺俁侽侽侽侽擭偱朞榓偡傞偲偄傢傟傞乯
丂丂偡側傢偪丄憂憿偐傜僲傾偺峖悈傑偱偺忋嬻偵偼丄岤偄悈忲婥偺憌偑偁偭偰丄侾師塅拡慄偵婲場偡傞扽慺侾係偺惗嶻検偑偼傞偐偵彮側偔丄俠14/俠12偺抣偼尰嵼傛傝傕偐側傝掅偄妱崌偱堦掕偟偰偄偨偲峫偊傜傟傞丅偟偨偑偭偰丄屆偄堚暔偵偮偄偰偼丄幚嵺傛傝傕偼傞偐偵挿偄擭寧偑宱偭偨傛偆偵應掕偝傟傞偺偱偁傞丅偨偲偊偽丄悽奅拞偵偁傞僲傾偺峖悈偺帪乮係侽侽侽擭慜乯偵暵偠崬傔傜傟偨壔愇偼丄栺俈枩擭慜偲應掕偝傟傞偺偱偁傞丅
丂丂乮仺丂抧憌擭戙偲僲傾偺峖悈乯
丂丂偙偺弶婜抣偼丄偄偔偮偐偺帋嶼偵傛偭偰尰嵼偺俆乣侾俆亾掱搙偲悇掕偝傟丄僌儔僼偵偡傞偲師偺傛偆偱偁傞丅
丂丂丂
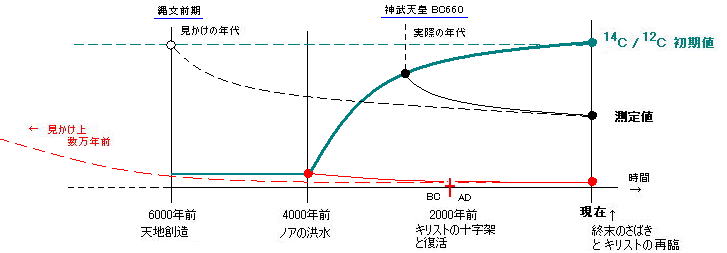
丂丂偡傞偲丄撽暥慜婜乮俇侽侽侽乣俆侽侽侽擭慜乯偲偄傢傟傞擭戙偼丄乮弶婜抣丒嬋慄偺愝掕偵傛偭偰偼乯恄晲揤峜偑懄埵偟偨偲偄傢傟傞崰乮俛俠俇俇侽擭崰乯偲廳側偭偰偟傑偆偺偱偁傞両乮俠14偺敿尭婜5730擭佮6000擭傛傝尰嵼偺敿暘丄偦偙偐傜俠14/俠12朞榓嬋慄偵栠偟偰岎傢傞強乯
丂丂丂撽暥憪憂婜乮侾枩擭慜丄偨偲偊偽墶恵夑丒壞搰奓捤偺僇僉妅丄栘扽偵傛傞應掕丗俋係侽侽擭丄峫屆妛揑偵偼崌傢側偄乯丅
丂丂丂撽暥慜婜乮俇侽侽侽亅俆侽侽侽擭慜丄奓捤丄搚嬼丄娵栘廙丄憰忺昳丒岡嬍丄奀柺忋徃丄婥壏偺忋徃乯偵偼侾侽枩偺恖偑偄偨偲尵傢傟丄暥壔偺拞怱偼搶擔杮偱偁偭偨丅
丂丂丂撽暥拞婜乮俆侽侽侽乣係侽侽侽擭慜丄嵟惙婜俀俇枩恖丄敳帟丄婥壏掅壓巒傑傞乯
丂丂丂撽暥屻婜乮係侽侽侽乣俁侽侽侽擭慜丄侾俇枩恖偵尭彮丄岎堈嫏楯柉丄惢墫丄僗僩乕儞僒乕僋儖乯
丂丂丂撽暥斢婜乮俁侽侽侽亅俀俁侽侽擭慜丄婥壏俀亷掅壓丄奀柺掅壓乮棽婲乯乮嫑夘椶巰柵乯偵傛傝俈-俉枩恖偵尭彮丄搶杒丗珞嫏丄杒嬨廈丒嬤婨丗撽暥悈揷乯
丂丂撽暥帪戙偺偡傋偰傪捠偟偰俛俠侾俆侽侽亅俛俠俀侽侽擭傎偳偱偁傠偆丅
丂丂丂丂憪憂婜丗丂壞搰奓捤乮恄撧愳乯丂丂慜婜丗丂撿杧奓捤乮恄撧愳乯丄扜寠廧嫃乮撧椙帪戙傑偱妋擣乯
丂丂丂丂拞婜丗丂塛嶳奓捤乮搶嫗)丂丂丂丂丂屻婜丗丂戝怷奓捤乮搶嫗乯丄壛慮棙奓捤乮愮梩丄崙撪嵟戝偺奓捤乯
丂丂丂丂斢婜丗丂婽儢壀堚愓乮惵怷丄婽儢壀幃搚婍乯丄斅晅堚愓乮暉壀丄嵟屆偺擾懞愓乯丄嵷敤堚愓乮嵅夑丄悈揷愓乯
丂丂偙偺偲偒傑偱偵丄搶傾僕傾宯偺恖乆乮杒奀摴偼杒傾僕傾宯乯偑擾峩傗揝婍丄暥帤側偳偺暥壔偑柍偄傑傑擔杮楍搰偺嬿乆偵傑偱廧傫偱偄偨丅斵傜偼丄撽暥堚愓偐傜椙偔抦傜傟偰偄傞傛偆偵丄扜寠幃廧嫃偵廧傒丄愇婍傪巊梡偟丄嫏楯丒庪椔丄僪儞僌儕側偳偺怉暔怘嵦庢丄埦丒僸僄偺嵧攟丒嶨崚偺庰丄暥帤側偟丄敳帟偺廗姷側偳偺惗妶偱丄暯嬒庻柦偼俀侽嵨偔傜偄偩偭偨丅堚崪偺條巕偐傜丄栱惗恖偲堘偭偰丄偁傑傝晹懓娫偺憟偄偼側偐偭偨傛偆偱偁傞偑丄塰梴幐挷偲昦婥偑尒傜傟傞丅
丂丂偟偨偑偭偰丄扨弮偵丄師偺壜擻惈偑峫偊傜傟傞丅俛俠俈俀侾擭傾僢僔儕傾曔廁偲側偭偨杒僀僗儔僄儖侾侽晹懓偺柉偼丄乽傾僢僔儕傾掗崙撪乿乮嘥楍侾俆丄侾俈乯傗乽儐乕僼儔僥僗偺斵曽乿乮嘥楍侾係乯丄傑偨丄墦偔傾僼僈僯僗僞儞杒晹乮乽僑僓儞愳乿乮嘥楌俆乯乯偵楢峴偝傟丄堦晹偼偦偺抧偵巆偭偨偑丄搶曽摬溮丒懢梲恄悞攓乮僄僛俉丗侾俇乯偺嫮偄僄僼儔僀儉偺柉偼丄僯僱償僃柵朣乮俛俠俇侾俀乯偺偡偱偵俆侽擭慜偵偼搶偺偼偢傟偺擔杮楍搰乮乽僔僯儉偺抧乮丠乯乿乮僀僓係俋丗侾俀乯乯偵傗偭偰偒偰丄抁婜娫偵撍慠掕廧偟偨偙偲偵側傞丅乭揤懛崀椪乭偺恄榖偲宯恾偼偒傢傔偰媽栺惞彂揑乮亖傾僽儔僴儉丄僀僒僋丄儎僐僽偺宯恾偲摨偠乯偱偁傞丅傑偨丄恄晲揤峜偑丄慏傗儔僶丄儔僋僟偵忔偭偰傗偭偰棃偨奊偑懚嵼偡傞丅
丂丂偙偺傛偆偵丄拞婜乣屻婜偺撽暥恖偼丄僀僗儔僄儖偺曔廁埲崀偲偄偆斾妑揑怴偟偄帪戙偵惗妶偟偰偄偨丅乮壜擻惈偼彮側偄偑乯傕偟丄恄晲揤峜偑幚嵼偟偰偄偨側傜偽丄偙偺帪婜偵丄懠偺搶傾僕傾宯偲嫟偵擔杮偵傗偭偰偒偰擔杮偵堏傝廧傫偩偼偢偱偁傞丅
丂丂乮俀乯丂屆戙恖偺俢俶俙暘愅偺摿挜丗
丂丂屆戙恖偺俢俶俙暘愅偼丄曐懚偺搙崌偄偵傛偭偰壜擻側傕偺偱傕偐側傝擄偟偔丄儈僩僐儞僪儕傾俢俶俙偑傗偭偲偱偁傝丄彫偝側抐曅偵夡傟偰偄傞側傜偽摼傜傟傞忣曬偼彮側偄丅傑偨丄倄堚揱巕偺忣曬偼暦偐傟側偄丅
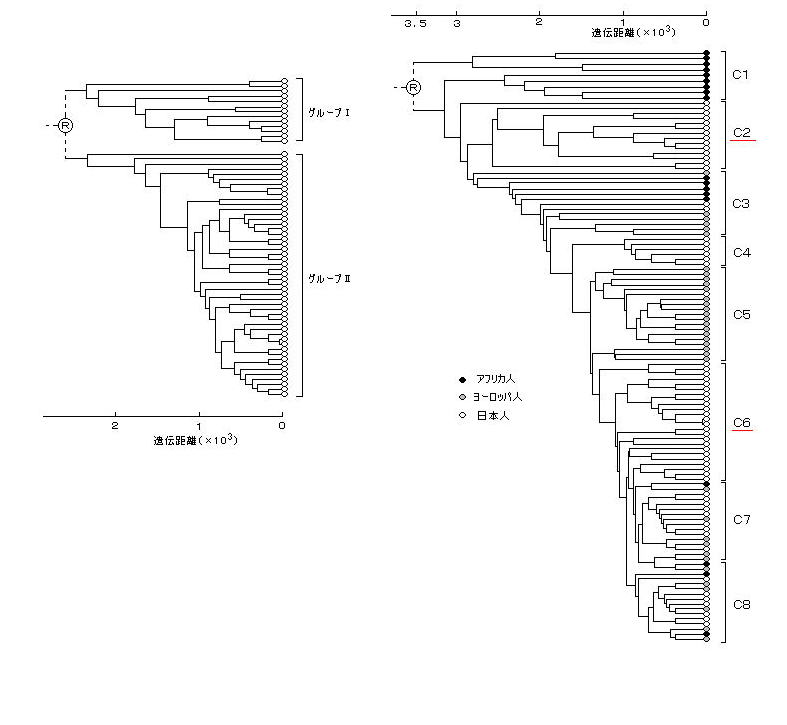
丂丂仏丂僔儀儕傾偺儅儞儌僗乮侾俋俋俋擭敪孈丄俁枩俆愮擭慜亖僲傾偺峖悈偺帪丗係侽侽侽擭慜丄搥搚憌亅侾俆乣亅俀侽亷乯偺俢俶俙偼丄慡懱偺俆暘偺侾偟偐巆偭偰偄側偐偭偨偺偱僋儘乕儞偼嶌傟側偐偭偨丅場傒偵丄恖娫偺僋儘乕儞乮栱惗恖偺僋儘乕儞側偳乯傪嶌傞偙偲偼乭僋儘乕儞朄乭乮俀侽侽侾擭俇寧巤峴乯偵傛偭偰嬛巭偝傟丄堘斀偡傞偲侾侽擭埲壓偺挦栶傑偨偼侾侽侽侽枩墌埲壓偺敱嬥偵側傞丅
丂丂曮棃巵偵傛傞丄撽暥恖偺儈僩僐儞僪儕傾俢俶俙偺弶婜偺曬崘偵傛傞撿曽宯愢乮僀儞僪僱僔傾乯偼椺奜揑乮俁専懱乯偱偁偭偨偑丄屻偵丄曮棃巵丄幝揷巵傜偑撽暥恖偺屄懱悢傪俆侽恖埲忋偵憹偟偰挷傋偨寢壥丄撽暥恖偲摨偠儈僩僐儞僪儕傾俢俶俙僴僾儘僞僀僾傪帩偮尰嵼偺恖乆偼搶傾僕傾側偳偵峀偔暘晍乮娯崙丄僇僓僼丄僂僀僌儖丄僽儕儎乕僩丒儌儞僑儖丄僀儞僪丄儓乕儘僢僷丄拞崙丒戜榩偺娍懓丄傾僼儕僇側偳乯偟偰偄偰丄偟偐傕丄栱惗恖俢俶俙傕偙偺抧棟揑暘晍僷僞乕儞偲偁傑傝堘傢側偄偲偄偆寢壥偱偁偭偨丅
丂丂丂丂丂丂丂丂丂丂

丂丂丂丂丂丂丂丂丂丂丂丂仏丂拞嵢堚愓偼嶉嬍導敀壀挰偵偁傝丄撽暥拞婜乣栱惗帪戙傑偱偺挿偄帪娫偺堚愓偑巆偭偰偄傞丅
丂丂丂丂丂丂丂丂丂丂丂仏仏丂係偺丄侾俈屄懱偵偮偄偰偼丄俶俫俲偑乭擔杮恖偺儖乕僣偼僽儕儎乕僩恖偱偁傞乭偲岆曬偟偨丅
丂丂偮傑傝丄乮應掕屄懱悢偼懡偔偼側偔丄堚愓偺抧堟傕曃偭偰偄傞偑丄乯撽暥帪戙丒栱惗帪戙弶婜偺帪揰偱偼丄偙偺寢壥偐傜扨弮偵丄擔杮偺乭愭廧柉乭偲偟偰偼傎偲傫偳搶傾僕傾宯偺恖偟偐偄側偐偭偨偙偲偵側傞丅
丂丂偡側傢偪丄倄愼怓懱俢俶俙偺俷宯摑乮搶傾僕傾宯乯偑杮搚擔杮偲壂撽偵丄俠俁宯摑乮杒傾僕傾宯乯偑杒奀摴偵丄偦傟偧傟愭廧柉偲偟偰懚嵼偟丄俢俀宯摑偑憹偊偩偟偨偺偼偦傟埲崀偲偄偆偙偲偵側傞丅乮偁傞偄偼丄嫃偰傕嫃廧抧偑曃偭偰偄偨傝丄恖岥偑彮側偐偭偨偺偐傕偟傟側偄丅乯
丂丂栱惗帪戙乣屆暛帪戙偵偐偗偰偼丄巟攝奒媺偵偁偭偨俢俀宯摑偺柉偑堦晇懡嵢惂偺偨傔媫憹偟丄儈僩僐儞僪儕傾俢俶俙乮曣宯乯偲倄愼怓懱俢俶俙乮晝宯乯偺傾僼儕僇丒拞搶宯偺斾棪偑傾儞僶儔儞僗偵側偭偨偲峫偊傜傟傞丅
丂丂傑偨丄僠儀僢僩宯儅僫僙懓偺枛遽偲峫偊傜傟傞乽恅巵乿堦懓偵偮偄偰傕丄係乣俆悽婭偺屆暛帪戙偵堏廧偟偰偒偨偺偱丄偙偺俢俀傪擔杮偵傕偨傜偟偨塭嬁偼戝偒偄偲峫偊傜傟傞丅
丂丂偄偢傟偵偟偰傕丄偙傟傜偺屆戙廤抍偼丄尰戙擔杮恖偲堎側傝丄搶傾僕傾宯偽偐傝偱偁傝丄傾僼儕僇丒拞搶宯側偳偼傎偲傫偳懚嵼偟偰偄側偄丅