日本人のDNA系統分析の特徴について
・・・・・ 日本人エフライム説の裏付け
2006 5/1
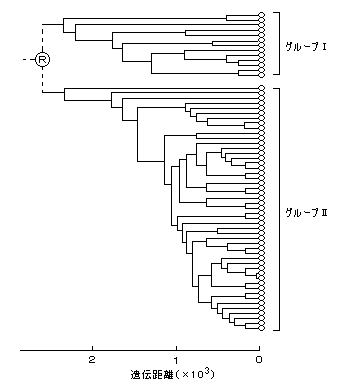
1. ミトコンドリアDNAの系統樹
ミトコンドリアDNAは、母系遺伝をするので、女性の祖先をたどった系図を調べることはできる。DNA配列が短期間に変化しやすいので分岐の時期が詳細に分かるという特徴がある。
宝来氏(故人)らによるミトコンドリアDNAの分析は、特に何の遺伝子も指定していない”Dループ領域”により多くの変異が観察されることから、この領域の482塩基の配列を2人づつ比較して、おのおのの配列間での塩基置換数を算出し、その数を遺伝距離(=横軸の長さ)とした。
((注) 当時、PCR法(下の注)(=DNA断片の増幅法)が発見されていなかったので、出産女性の後産が比較的多量に得られるミトコンドリアDNAの試料であり、これを用いて母系遺伝が先に研究された。)
その結果、日本人(静岡県・三島、62名)のミトコンドリアDNAの遺伝距離(横棒の長さ=塩基の入れ替えの数)によって作成された系統樹は、大きく2グループに分かれた。これを、グループⅠ、Ⅱとする。(右図)
また、同氏による、日本の地域(青森、静岡、沖縄)別のミトコンドリアDNAのグループは下図のように、北ほどグループⅠの割合が多くなった。(三地域の平均16%)
女性が男性に比べ定着後大きな地域的移動をしなかったのであれば、母系遺伝をするミトコンドリアDNAではこのような
地域に特異的なタイプが残っていると考えられる。
地域
|
個体数
(人) |
グループⅠの頻度(%) |
塩基多様度(%) |
共通タイプ(%) |
| 青森 |
61 |
28 |
0.26 |
33 |
| 静岡 |
116 |
18 |
0.26 |
39 |
| 沖縄 |
82 |
5 |
0.16 |
33 |
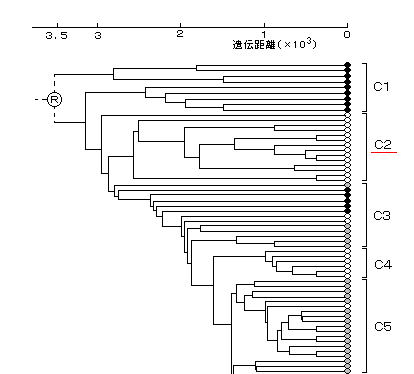
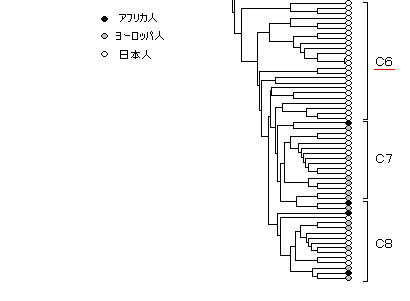
さらに、世界的に見たこの日本人のミトコンドリアDNAの2重構造の位置付けを知るために、アフリカ人、ヨーロッパ人と、日本人の間で系統関係を分析すると非常に興味深い結果が現れる。(下図) (その他のアジア人は入っていない)
宝来氏は、当初、縄文人と弥生人の区別を実証する目的でこの研究を行なったが、その分岐が世界3大人種を区別するほどの根元のほうにあるグループ
C2 が存在することを説明することはできない。これは、後の研究により、縄文人と弥生人のDNAは共に東アジアに広く分布するアジア系で、どちらが北でどちらが南から来たかという地域差がないことが分かってきた。従来行なわれてきた骨の形状などの”形態学”は当てにならないのである。(→
(参照)の3)
C2 の日本人グループに注目すると、その分岐点はヨーロッパ人よりもはるかに根元の アフリカ人グループの間にあるので、この日本人のミトコンドリアDNAは、聖書による3大祖先 ・・・ ハム、セム、ヤペテ ・・・ のうちの、セム系に相当すると考えられる。
また、C4、C6 、すなわち、日本人のアジア人種的なグループが、ヨーロッパ人にきわめて近い所で分岐している(=外見はどうであれ、DNAではきわめて近縁である)ので、ヨーロッパ人種とアジア人種が、残りのヤペテを同一の祖先に持つと思われる。
したがって、C2 が上のグループⅠに、C4、C6 がグループⅡに、それぞれ相当すると考えられる。(→ (参考)の4)
ヨセフの子エフライムの母は、エジプトの祭司ポティ・フェラの娘アセナテであった。(=ハム系)(創41:50-52)(*当時のエジプトにはまだアレクサンダー遠征によるギリシャ人の血が入っていない) しかし、ヨセフの子供に娘がいたとは書かれていないので、母系遺伝のミトコンドリアDNAは早くもここで切れている。したがって、系図のしるしとしては、セム系であるらしいことしか分からないのである。
エフライムには、ハム系の遺伝子が常染色体、および、X染色体上の遺伝子として半分混じっているはずである。(エジプトの祭司の血筋なので、しるしと不思議のカリスマ信仰に反応しやすい?) (現在のユダヤ人のユダ族にも途中でハム系のラハブが入っている。)
東北地方にはエフライムやユダの末裔が多いのではないかと考えられるが、日本全体にかなりの割合(平均16%)で存在していることになる。カナンの地を征服するため戦ったヌンの子ヨシュアはエフライム族のかしらであった。(民13:8、:16)
ヨセフ族、エフライム族にはリバイバルにかかわる特別な祝福が預言されている。(創49:22-26、申33:13-17)
2. Y染色体DNAの系統樹
PCR法が発見されてから本格的に調べられた Y染色体のDNAは 父系遺伝する。これは男系子孫にのみ伝わっていくので歴史的な調査に用いられ、(男系が続いていたならば)人種のルーツの祖先を正確に辿っていくことができる。
ただし、Y染色体にあるDNAのうち、機能を持つ遺伝子はきわめて少ないので、初代の人の体型・体質、能力や性格などの遺伝子の特徴が代々その子孫に伝えられることはなく、あくまでも過去の歴史のしるしである。(大部分の遺伝子は、父と母から伝えられる”常染色体”と、母親のみから伝えられる”X染色体”上にあるので、その具体的に現れる遺伝子セットの特徴は、数代で立ち消えになる。)
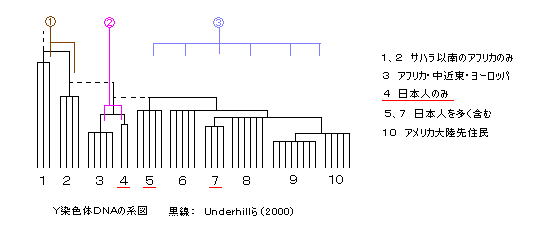
世界の各地のY染色体DNAの多数タイプは、図の10系統に分類され、歴史的に父系が続いてきた家系のグループについては存続し、聖書的な観点からは、おおよそ ①、②、③がそれぞれ、ハム系、セム系、ヤペテ系であると考えられる。
4 の系統は日本人のみ(東南アジアが1人だけいるが)であることから、この特別な系統はセム系(神武天皇(エフライム)、秦氏(ユダ?)などのユダヤ系)ではないかと思われる。4 は分岐の位置から、明らかにハム系(1、2)とは別のグループであり、3
のアフリカ中心、中近東、ヨーロッパの一部、に近い。
3 にはアジア人は全く含まれていない。したがって、4 のグループは、完全に3 から分離し、はるか西のほうからシルクロードを通って そのまま日本まで父系を保ちつつ独自に移動してきたと考えられる。(途中でどのような女性をめとったかは問題ではない。)
マナセ族は一度男子が切れている(民26:33、27:1-)が、主の命令により、彼女たちの父の部族の氏族にとついでいる(民36:6)ので父系はつながっている。
アフリカ南部のレンバ族の祭司は世襲制であり、Y染色体のDNAがよく保存され、ユダのレビ族と同じアロンの子孫の特徴が、この外見上全く似ていないレンバ族にはっきり認められた。(”ネイチャー”:1997 1)
(参考文献)
・ 「DNAから見た日本人」、斎藤、ちくま新書、2005
3 ・ 「DNA人類進化学」、宝来、岩波科学ライブラリー、1997
(・ 「日本人の来た道」、池田、朝日選書、1998 ・ 「日本古代史の謎」、自由国民社)
(参考)
(1) DNAと突然変異について:
ヒトのゲノム(46本)は、22対(44本)の常染色体と1対の性染色体(2本、男XY・女XX)に分かれて収められている。各染色体は一本の長いDNAのひもで成り立っていて、その長さは 1m強で、塩基数にして 約30億個ある。ヒトゲノムの中には、3万個程度の遺伝子が入っているが、そのうち遺伝情報としての意味を持っているのは全体の5%程度である。(2003年の春にヒトゲノムの99%が解読されたが、それがどのような意味を持つかは大部分が解明されていない。体細胞の数は60兆個あるので、一人のヒトのすべてのDNAを一直線につなげると60兆m
= 太陽系の直系位 になる。) ヒトの遺伝子の95%以上は常染色体上にある。
DNA分子の自己複製の時、塩基の”置換”、”挿入”、”欠失”などの間違いをまれに犯す。特に、”CACACA・・・”、”GACGAC・・・”のような2あるいは3塩基の繰り返しがある所で、その繰り返しの回数を変化させる変異の発生率が高い。ヒトゲノムの95%以上を占める 意味を持たないDNA(”がらくたDNA”)の部分は、機能を持つDNAの部分よりも突然変異の割合がはるかに多い。(中立突然変異:生存に有利でも不利でもない変異の説明)
突然変異は通常、1個体から始まる。仮に、2500年前に突然変異した1人の人がいると、当時の日本人の人口30万人は現在1億2000万人であるから400倍、つまり2500年間で400人しかならないことになる。ある遺伝子を持つ人が日本に100万人いたとすると、それ以前に変異を受けていたことになる。
ミトコンドリアDNAの塩基置換速度(=”進化論”的立場で言うところの”進化速度”)は、100万年当たり少なくとも1%程度と推定されている。
現在のヒトの間のDNAの違いは、推定で約0.07%maxで、チンパンジーとの間の差異 1.2%よりもかなり小さい。他人は自分のほとんどコピーであるといえる。 ウィルソンの「単一起源説」(”第2の出アフリカ説”、(1987))によると、53人のミトコンドリアDNAの塩基すべての比較により、15―20万年前:東アフリカにミトコンドリア・イブが出現、10万年前:アフリカ人が分岐、5万年前:ヨーロッパ人、アジア人が分岐したことになっている。
・・・・・ ただし、進化論に必須の長い年月の想定は、ノアの洪水(4000年前)を仮定すると、洪水により炭素14の量が激増したことから、大幅に修正を求められる点である。これは進化というより、遺伝子にキズがついたという程度で、近親結婚で奇形が発生するレベルであり、サルから数%も変化するには余りにも時間が足りなすぎる。まして、機能を持つ遺伝子が存在する所のDNAが変異することはほとんどあり得ない。ただし、ヤペテからヨーロッパ系とアジア系の人種が分かれたのは突然変異によるのかもしれない。
(2) ミトコンドリアDNAと Y染色体DNAの遺伝について:
Y染色体DNAは 父親から男の子のみに伝わり(男→男の子→男の孫→・・・)、ミトコンドリアDNAは、(受精卵の中で父親からの精子にあるミトコンドリアDNAは破壊されてしまうことが知られている) 母親から女の子にのみに伝わる(女→女の子→女の孫→・・・)。したがって、たとえば父親に女の子しかいないと、その Y遺伝子はそこで途絶えることになり、祖先のDNAの追跡はできない。もっとも、同じ Y染色体をもつ近縁の者との間の子には、このしるしを伝えることができる。同様に、母方の家系の血を引く女性をめとった場合もミトコンドリアDNAは引き継がれる。
* かつてモンゴル帝国を築いたチンギス・ハンではないかと推察される Y染色体DNAのハプロタイプ(複数の遺伝子座の対立遺伝子の組合せ)が、異常に高い頻度でモンゴルを含む中央アジアの集団で存在している。モンゴルは長い間男系子孫が王位継承権を持ち、また各地で多くの女性と交渉を持ち多くの子孫を残したといわれる。ただし、Y染色体上における機能的な遺伝子はほとんど無く、交配によって再び 政治や軍事にすぐれた英雄が現れることはない。 ちなみに、このハプロタイプは、日本では見つかっていない。(したがって、源義経がチンギス・ハンになったという説はうそである。)
(3) 古代DNAの調査結果:
宝来氏は、縄文人のミトコンドリアDNAが東南アジア人のミトコンドリアDNAに類似していることから、従来の形態学による縄文人の東アジア南方起源説と弥生人の北方移住説を支持した。しかし、これは例外的(3検体)であり、後に縄文人の個体数を50人以上に増して調べた結果、縄文人と同じDNAハプロタイプを持つ現在の人々は東アジアに広く分布(日本列島、朝鮮半島、中国、ブリアート人)していて、しかも、弥生人DNAもこの地理的分布パターンとあまり違わないという結果であった。つまり、宝来らによる日本人のDNAの2重構造は、定説となっていた縄文人と弥生人という2重構造ではない。(by.篠田謙一)
長い間研究されてきた古代人骨による「形態学」は、それほど信憑性のあるものではないことが明らかになった。たとえば頭蓋骨の”短頭化現象”は、日本のみならず世界的な傾向である。また、明治以降の150年の間に日本人の身長が著しく伸びた。
すなわち、形態は、遺伝要因のほかに環境要因(栄養条件、気候条件など)によって大きく異なるのである。ちなみに、短頭・長身・細目の朝鮮系の渡来人は近畿地方に多く、歴史的に根拠があるとされてきた。
大陸からの渡来が4回に分けてあったが、朝鮮人を含む東アジア6集団(韓国人、モンゴル人、本土日本人、チベット人、アイヌ人、沖縄人)はDNAはそれほど大差は無い。 尚、骨格や体節構造を支配するDNAは現在探求中であり、未だよく分かっていない。
(DNA操作により巨大化する”巨人”は作られ得るか?「ネフィリム」、「アナク人」)
* 大陸からの渡来人は、これ以前を除いて、① BC2-3c: 弥生人、北九州に朝鮮・中国系の遺跡・遺物、②
5c:倭の五王の時代に朝鮮系、製鉄技術、鉄製農具、灌漑・土木、③ 5c後半-6c:
技術者集団→大和朝廷を強める、 ④ 7c後半:百済の亡命者、仏教・仏像伝来、となる。
特に、②あるいは③の時期に、秦(はた)氏一族が5c後半に来朝し、その後、最大の古代豪族になった。秦氏は 弓月君(ゆづきのきみ)と称したが、秦の始皇帝の子孫という説には根拠が無く、中国西端の弓月(クンユエ)出身であり、ユダヤ系(しかも、ネストリウス派の東方キリスト教徒)であったらしい。 5cに、彼らは、新羅・百済と同盟して 北からの高句麗に対抗しようという倭の朝鮮外交にかかわって、大和朝廷は”秦伴造(はたのとものみやつこ)”という官人として受け入れられた。(農業、機織、銅の鋳造(和同開珎)、木工などにより、殖産的・田園的豪族として、6cの欽明朝の頃 戸籍につけられた数は”秦人戸数 7053”にものぼった(美濃・伊勢・尾張、越前・越中、播磨・美作・備前・備中、讃岐・伊予、豊前・筑前)。また、後には 太秦(うずまさ)に本拠を移した。6c末-7c初には、秦造河勝(はたのみやつこかわかつ)は 聖徳太子(厩戸(うまやど)皇子)の側近として、物部氏との戦いの軍政人、新羅使節の迎接官として活躍した。
(4) アジアとヨーロッパの共通祖先:
中国の山東半島の付け根にある (りんし)という地域の遺跡にある、同一地域の異なる時代(前漢末期・2000年前、春秋戦国時代中期・2500年前)の人骨資料のミトコンドリアDNA分析を行なった結果、前者は東アジアではなく中央アジア集団に近く、後者はさらにヨーロッパ集団に近いことが分かった。古代中国ではこのように人種が錯綜して、今後の中国の古代史に新しい解釈を迫る可能性がある。
(りんし)という地域の遺跡にある、同一地域の異なる時代(前漢末期・2000年前、春秋戦国時代中期・2500年前)の人骨資料のミトコンドリアDNA分析を行なった結果、前者は東アジアではなく中央アジア集団に近く、後者はさらにヨーロッパ集団に近いことが分かった。古代中国ではこのように人種が錯綜して、今後の中国の古代史に新しい解釈を迫る可能性がある。
ミトコンドリアDNAの分岐からは、アジア6集団の近縁性に次いで、外見上は全く異なるヨーロッパ人との近縁性がある。これはアフリカ人やセム系よりもはるかに近縁である。
ちなみに、中国南部の苗族(ミャオ族)にはヤペテ系の祖先をもつ伝承がある。
(5) 血液たんぱく質の分析:
1960年代、でんぷんゲル電気泳動法が普及し、血液中のたんぱく質のアミノ酸配列がDNAの塩基配列に対応することを利用して、常、X染色体上の遺伝子のDNA配列が比較調査された。この場合、ミトコンドリアやY染色体のように1組の祖先に行き着くもの(系統樹)ではなく、遺伝子は混合してしまうから、集団間の相対的な近縁性の比較のみである。
その結果、分岐の程度は、ヨーロッパ-北インド・イラン-北米・南米-東アジア-東南アジアの順にほぼ等間隔でグループが分かれ、それぞれの間の遺伝距離は、アフリカとの分岐に比べるとそれほど遠くなかった。ただし、東アジアの6集団は非常に接近していた。(1997)
* 日本では、俗に”ABO式血液型の性格判断”が言われてきたが、ABO式血液型物質が発現するのは血液系や上皮組織の細胞であり、性格に影響を与える脳神経系ではないから、血液型と性格との相関は無いと考えられる。
* 「DNAが解き明かす日本人の系譜」、崎谷満、勉誠出版、2005
8 によると、
”Y染色多型体の分析による最新の結果では、旧石器時代(?)にシベリア経由で北海道に来たC3系統、縄文時代(?)に華北・朝鮮半島経由で来たD2系統、弥生時代に同じく華北・朝鮮半島経由で来たO2b、O3系統が、(沖縄を含む)日本列島のヒト集団を形成し、南方から沖縄経由で来た痕跡はごくわずかである。
アフリカ(?)をあとにして広がった人類は大きく3つのグループに分かれたが、日本列島にはその3つのグループのすべてが集まっていて、これは全世界的に見て他に類の無い特徴である”、とある。
・・・・・ ヨセフは、エジプトの祭司ポティ・フェラの娘アセナテと結婚してエフライムとマナセが生まれた。(創41:50) Y遺伝子は父系遺伝する。 したがって、日本人には、一般の東アジア系(ヤペテ系: 縄文系+弥生系といわれたグループ)に、ヨセフのユダヤ系(セム系)とエジプト人(ハム系)のDNAが入っていることとよく合っている。
(2006 12/1追加)
トップへ戻る