ミトコンドリアDNA分析(母系遺伝)では、日本人を構成する系統は大体2種類に分けられ、しかもその一つはアジア系ではなく、むしろアフリカ系に近い、アジア系とは非常に異なるDNAが含まれていることが明らかになった。(Y染色体の遺伝子がほとんど機能を持たない形式的な”しるし”であるのに対し、常・X染色体の遺伝子は、人種の形質に大きな影響を与える。)( → ミトコンドリアDNAの系統樹 )
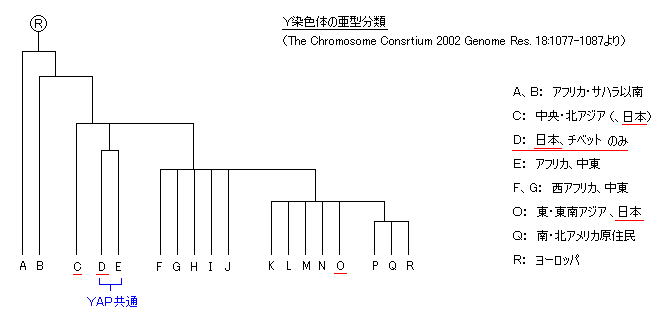
一方、父系遺伝するY遺伝子のDNAには、さらに明らかな大きな特徴があることが判明している。旧石器時代(?)にシベリア経由で北海道に来たC3系統(中央・北アジア系)、縄文時代(?)に華北・朝鮮半島経由で来たD2系統、弥生時代に同じく華北・朝鮮半島経由で来たO2b、O3系統(東アジア系)が、北海道(アイヌ)から沖縄島までの日本列島のヒト集団を形成したと考えられる。
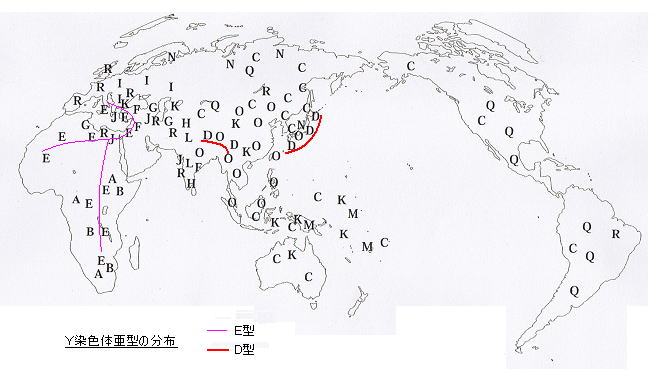
日本列島のDNAは、このD、O系統でほとんどを占め、D2の割合は半分弱にも達する。D2は日本列島にまんべんなく分布し、特にアイヌ人には9割近くも集積している。(本土の山間部にもやや多い。) 大陸からの弥生人が流入した地点の九州では若干少ない。沖縄島では若干多いが、それよりも南の八重山諸島にはD2は無い。
このD2系統には、特徴的な変異:「YAP」(ヤップ)とよばれる挿入部分(300塩基)をもち、これは D、E系統のみに含まれ他には存在しない。E系統は、北・西アフリカ、ナイル川沿い、地中海沿岸などのアフリカ・中東に広く分布し、ナイジェリアにその祖型(DE*)があるという報告がある。そこからかなり昔に分岐した D系統は、現在、日本とチベットのみに著しい集積が見られ、他の地域では非常に少ないか全く無い。
東アジアで比較的多い朝鮮でさえ2〜4%と少ないということは、絶滅されたというよりは、その中心集団が日本列島に移動しきったためと考えられる。
また、イスラエルの失われた10部族の調査機関、『アミシャブ』によると、このチベット近辺には、シンルン族(メナシュ(=マナセ)族)、カレン族、チャンミン族という10部族、特に、マナセ族の宗教・伝統を今もなお継承している民族が存在していることが明らかにされている。また、アミシャブは日本について、文化的考察から、日本人と失われた10支族の間に何らかのつながりがあることを否定できないとしている。
唯一、特別な動きをしたヨセフの記録は、創世記41:50−52に書かれてある。ヨセフの妻 エジプトのオンの祭司ポティ・フェラの娘アセナテからそのままエフライム、マナセに入ったが、母系遺伝するミトコンドリアDNAの分析ではよく説明していると見ることができた。
しかし、Y染色体の遺伝子は単純に父系遺伝するので、YAP他のD系統の特徴が、あたかも先祖の父親がハム系のように見えるのである。これは、5.で考察する。

マナセ族の末裔の居住地(by.アミシャブ):
